The Memory Field: Could Quantum Biology Involve Accessing Information Stored in Space Itself?
TL;DR
What if memories aren't "stored" like files inside your brain? Emerging evidence suggests a radical alternative: experiences leave patterns in the fabric of space itself, and subcellular structures inside neurons act like resonant dials that retune those patterns on demand. Synapses help you find the right station; the broadcast lives in spacetime.
The Storage Paradox: What Hyperthymesia Reveals
Is there any reason to suspect that the conventional model purporting to explain cognitive faculties via neuronal correlates is incomplete or even inaccurate? The conventional story seems to explain the situation logically and matches certain observable data: neurons that wire together fire together, which is seen in a process called neuronal plasticity and ostensibly the patterns formed by networked neurons are memory traces. In this view, connections between neurons, called synapses, are like digital bits that record an on/off state, and that is how information is encoded in the brain. This is called the neurocomputational model.
But then, some people exist who severely challenge this computationalist paradigm—individuals with Highly Superior Autobiographical Memory (HSAM). As we will see, the observables of this condition seriously challenge certain aspects of the neurocomputational model, because there just aren't enough "bytes" available via synapses to account for HSAM.

You've probably heard of amnesia, which is a total loss of memory. There is an opposite phenomena in which individuals cannot forget some biographical information, called Hyperthymesia. Also referred to as Highly Superior Autobiographical Memory (HSAM)—is a rare ability where everyday experiences are recalled with striking, movie-like detail. Ask someone with HSAM about a random date years ago and they can often tell you what they were doing, what the weather was like, maybe even what they ate or wore—without special tricks or rehearsal. These memories arrive quickly, feel richly textured, and are tied to exact calendar dates.
A few clarifications:
· It’s not “photographic memory.” HSAM is specifically about personal, day-by-day life events, not instantly memorizing phone books or textbooks.
· It’s spontaneous, not staged. The recall comes naturally rather than from trained mnemonic methods.
· It’s encyclopedic over time. What stands out is the sheer volume of accurate, date-anchored memories across many years.
Why this matters here: if we treat synapses like a digital storage device with a fixed number of bits, the amount of information HSAM seems to retain—high-fidelity, calendar-indexed memories across decades—overwhelms the budget. This presents a clear observation-theory mismatch: if synaptic networks truly function as biological hard drives, the storage demands of hyperthymetic recall should exceed the physical constraints of neural architecture by orders of magnitude—suggesting we need to reconsider the foundational assumptions of the computational memory model.
The problem with the brain-as-hard-drive story
Conventional neuroscience reduces the neuron to what anesthesiologist and consciousness researcher Stuart Hameroff calls a "cartoon neuron"—a simple on/off logic gate that ignores the rich subcellular machinery within. Reducing the neuron to a simple logic gate makes the neurocomputational model easy to understand: the synapse is a bit and lots of bits enable computation and computation is necessary and sufficient for consciousness (a view called computational functionalism [1]). However, such a simplistic model misses the actual functionality of the neuron and synapse, which operates as a multi-scale cybernetic resonant feedback system—a dynamic network of EM potentials and molecular interactions that can coordinate across multiple temporal and spatial scales, rather than merely switching on or off like a digital gate. To illustrate why this distinction matters for memory storage, we can examine the capacity constraints that emerge when we take the computational model to its logical conclusion.

Take the most generous “cartoon neuron” estimate: treat each synapse like a one-bit switch and multiply synapses per neuron by total neurons. You get on the order of a quadrillion bytes (~1 petabyte) of capacity for the whole brain. That sounds big—until you compare it to lived experience.
A petabyte holds only ~15.5 days of continuous, uncompressed 4K video. People with hyperthymesia report essentially movie-like recall of everyday life across decades. Translating that into the same video analogy, maintaining 60 years of richly detailed daily recall would demand roughly ~1,000 petabytes (≈1 exabyte)—about a thousand times the cartoon model’s upper bound.
And the picture tightens further if you confine “storage” to the commonly cited memory hub, called the hippocampus (with some memory storage distributed in the cortical layers). The hippocampus is ~0.5% of brain volume; on the cartoon model, that’s ~5 terabytes—about the size of a good external drive. Useful, sure. But not nearly enough for a lifetime of cinematic recall.
Bottom line: synapses are essential, but their role fits far better as indexes and tuning knobs—pointers that help you find the memory—rather than as the vault that literally stores it.
Space as a memory medium
Modern physics increasingly treats spacetime as an active, fine-grained substrate—more like a dynamic medium than empty nothingness. This can be seen in theories from Qauntum Memory Matrix [4]—in which space–time itself acts as a dynamic quantum information reservoir, with quantum imprints encoding information about quantum states and interactions directly into the fabric of space–time at the Planck scale—to the holographic principle itself developed by Gerard t'Hooft, Leonard Susskind, and other preeminent physicists. On this view, every interaction, be it the passage of a particle or even the influence of a force such as electromagnetism or gravity, leaves a quantum imprint into the fine-scale structure of spacetime. This, of course, includes the motions and interactions of the animate matter comprising living systems, which will also leave faint yet durable quantum imprints. These modifications can be richly complex for biological structures, like the subcellular cytoarchitectonics of neurons, generating veritable micro-scale signatures that record indelible imprints in spacetime.
Memory, then, isn't just wiring in the head; it's a pattern in space that can be re-accessed through resonant coupling to these persistent spacetime signatures.
Recollection becomes less like opening a saved file and more like re-tuning a resonator to a pattern that still exists in the underlying field.
The cellular “stylus”: microtubules and friends
Inside your neurons is an intricate scaffolding—microtubules—built from protein subunits that host ring-shaped molecular groups with lively electron clouds. In ordered arrays, these rings can share energy, phase-lock, and coordinate like miniature antennae. In short: they’re natural subcellular resonators.
Our proposal: networks of these resonators entangle with each other and with the fine structure of space. When a memory is formed, the brain’s resonant state imprints a pattern in the field. When you remember, microtubule networks slip back into the same resonant configuration, coupling to the original pattern. Synapses help steer the system to the right “address,” but the content is recovered through resonant access.
Entanglement as a bridge (in plain language)
“Quantum entanglement” links things so tightly that changing one instantly shapes the state of the other, no matter the distance—without sending a signal. A growing line of theory suggests these links are tied to actual connectivity in spacetime (often summarized by the slogan ER = EPR). If biology can sustain robust entanglement at the molecular scale—especially in ordered structures like microtubules—then the brain already has the right kind of hardware to plug into the spacetime network.
Think of spacetime as a vast vinyl record and microtubules as the stylus. Daily life etches grooves; later, the stylus rides the same groove and the music plays again. The song isn’t inside the stylus—it’s in the groove.
Why some people recall so much (Hyperthymesia)
A helpful mechanism comes from Hameroff and colleagues: the enzyme CaMKII (Figure 1)—a key player in synaptic plasticity and long-term potentiation [2]—can phosphorylate specific molecular “registers,” acting as an indexing system that flags where and how to tune a memory [3]. In typical brains, this indexing is constantly trimmed (a process called dephosphorylation) and updated as experiences are integrated—helpful for avoiding overload.
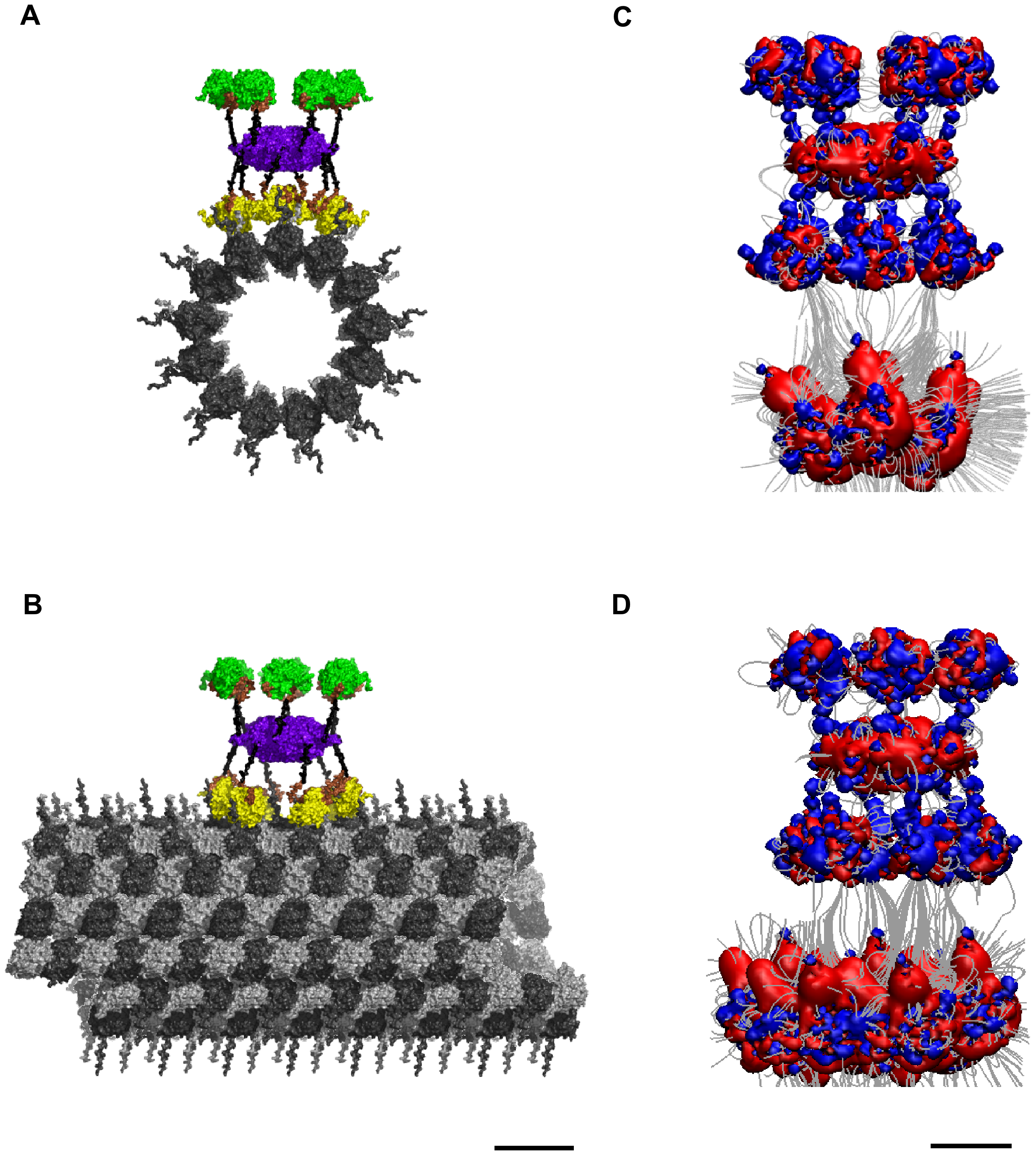
Figure 1. CaMKII indexing on microtubules creates phosphorylation registers.
(A) Top view of a CaMKII holoenzyme poised above the microtubule (MT) outer surface. The hub (violet) organizes kinase modules (green) via flexible linkers (brown), positioning them to reach the MT lattice. (B) Side view of CaMKII docked on the MT; catalytic heads phosphorylate exposed side chains on the MT surface (yellow), establishing persistent molecular registers that can serve as synaptic indexes. (C–D) Field visualization of the catalytic footprint: red/blue lobes indicate the spatial domain over which kinase heads can act when the holoenzyme is engaged with the lattice, highlighting how clustered phosphorylation can write discrete, addressable tags along protofilaments. MT: dark gray. Scale bars as indicated. Image from: T. J. A. Craddock, J. A. Tuszynski, and S. Hameroff, “Cytoskeletal signaling: is memory encoded in microtubule lattices by CaMKII phosphorylation?,” PLoS Comput Biol, vol. 8, no. 3, p. e1002421, 2012, doi: 10.1371/journal.pcbi.1002421.
In hyperthymesia, the hypothesis is that this pruning is reduced or effectively irreversible for certain autobiographical traces. Imagine an enzyme responsible for surveillance and updating of "phosphorylation registeries" by CaMKII, but it has a mutation that significantly limits its capabilities, leading to an inability to maintain only salient tags. The result: exceptionally stable, highly specific indexes that make “tuning back in” almost effortless and extraordinarily vivid.
Note: This means that this resonant-access model is consistent with most aspects of neurobiological theory of consciousness [5] as well as long-term potentiation being a primary mechanism underlying learning and memory [6], it just posits that the correlates of these cognitive and sentient capacities are involving subcellular systems and quantum gravitational dynamics.
The gravitational wave memory effect: how space keeps score
Here’s a physical channel for the imprint itself. Work by ISF researchers has demonstrated that every particle can be modeled as a tiny black-hole/wormhole system whose dynamics continuously generate gravitational waves [7, 8]. When matter moves, accelerates, or changes configuration, it doesn’t just pass through space; it writes to it.
Gravitational waves can leave a lasting offset in spacetime called the gravitational wave memory effect—a permanent “afterglow” of the event in the geometry itself. In this picture, life’s ongoing activity etches a distributed record into spacetime. Because ER = EPR links entanglement with spacetime connectivity, the entanglement structure can reach back across space to the coordinates where experiences were etched, letting resonant biological interfaces pull those patterns forward into awareness.
In short: the brain tunes; spacetime remembers.
What this doesn’t claim
· It doesn’t say neurons are unimportant. On the contrary: neural activity choreographs the tuning process.
· It doesn’t invoke sci-fi time travel. In plain physics terms, recall is re-entering an existing entangled pattern—no rule-breaking required.
· It doesn’t deny plasticity. Synaptic change remains vital; it just shifts role—from warehouse to wayfinding.
· It doesn’t exclude other subcellular organelles, like mitochondria, that play central and ineluctable roles in cognition.
Why it matters
· Better fit to experience. A resonant-access model makes sense of lifetimes of richly detailed memory and the sudden wholeness of certain recollections.
· New strategies for brain health. If memory loss involves failed tuning rather than missing “files,” we can target subcellular resonance—opening possibilities beyond today’s approaches.
· Blueprints for new technology. Nature is utilizing non-computational information access through resonance, entanglement, and geometrical memory in spacetime. With an understanding of this mechanism we can engineer entirely new information processing and storage systems that mimic biological quantum-gravitational memory mechanisms. These technologies could include:
1. Quantum resonance memory devices that store information through entangled field patterns rather than traditional digital encoding
2. Biomimetic information retrieval systems that access data through vibrational and quantum coherence principles
3. Consciousness Interface Systems that implement quantum gravitational resonant architectures with cross-scale feedback
By recognizing memory as a fundamental property of spacetime, we open pathways to technological paradigms that transcend current computational limitations.
If you’re interested to learn more about this research, check out the full presentation by following the link below”
William Brown Presents at the Science of Consciousness Conference, Barcelona 2025, Following Sir Roger Penrose
July 7, 2025 - Plenary-3 - ‘Consciousness and Reality':
Sir Roger Penrose (R) Classical and Quantum Realities are Not Quite the Same; How About Conscious Reality?
Start time 47:57- William Brown, Instantaneous Memory Accession via Quantum Geometrodynamic Networks
Start time 1:23:53- James Tagg, Are quantum computers already conscious?
References
1. A. K. Seth, “Conscious artificial intelligence and biological naturalism,” Behavioral and Brain Sciences, pp. 1–42, Apr. 2025, doi: 10.1017/S0140525X25000032.
2. W. Tao et al., “Synaptic memory requires CaMKII,” eLife, vol. 10, p. e60360, 2021, doi: 10.7554/eLife.60360.
3. T. J. A. Craddock, J. A. Tuszynski, and S. Hameroff, “Cytoskeletal signaling: is memory encoded in microtubule lattices by CaMKII phosphorylation?,” PLoS Comput Biol, vol. 8, no. 3, p. e1002421, 2012, doi: 10.1371/journal.pcbi.1002421.
4. F. Neukart, R. Brasher, and E. Marx, “The Quantum Memory Matrix: A Unified Framework for the Black Hole Information Paradox,” Entropy, vol. 26, no. 12, p. 1039, Dec. 2024, doi: 10.3390/e26121039.
5. J. Aru, M. Suzuki, and M. E. Larkum, “Cellular Mechanisms of Conscious Processing,” Trends in Cognitive Sciences, vol. 24, no. 10, pp. 814–825, Oct. 2020, doi: 10.1016/j.tics.2020.07.006.
6. R. Yasuda, Y. Hayashi, and J. W. Hell, “CaMKII: a central molecular organizer of synaptic plasticity, learning and memory,” Nat Rev Neurosci, vol. 23, no. 11, pp. 666–682, Nov. 2022, doi: 10.1038/s41583-022-00624-2.
7. N. Haramein, C. Guermonprez, and O. Alirol, “The Origin of Mass and the Nature of Gravity,” Sep. 2023, doi: 10.5281/zenodo.8381115.
8. N. Haramein, O. Alirol, and C. Guermonprez, “Extending Einstein-Rosen’s Geometric Vision : Vacuum Fluctuations-Induced Curvature as the Source of Mass, Gravity and Nuclear Confinement,” Sep. 23, 2025, Preprints: 2025091835. doi: 10.20944/preprints202509.1835.v1.